Un estudio español en Nature replantea el origen de nuestras células como una historia de alianzas microbianas
El origen y el proceso por el que surgieron las células eucariotas es aún una de las grandes preguntas de la biología, con las teorías de Lynn Margulis sobre la incorporación de una bacteria que luego sería la mitocondria como gran punto de inflexión. Ahora, una investigación española realizada desde el IRB Barcelona y el Barcelona Supercomputing Center replantea esta visión. Sin negar el papel de la mitocondria, apunta a que el proceso fue más largo y complejo de lo que se pensaba, prolongándose durante cientos de miles de años. Al menos otras dos bacterias diferentes contribuyeron al desarrollo de las células eucariotas y virus gigantes parecen haber actuado como vehículos de transferencia genética. Los resultados, que se publican en Nature, sugieren un proceso mucho más dilatado y gradual de intercambios entre microorganismos.
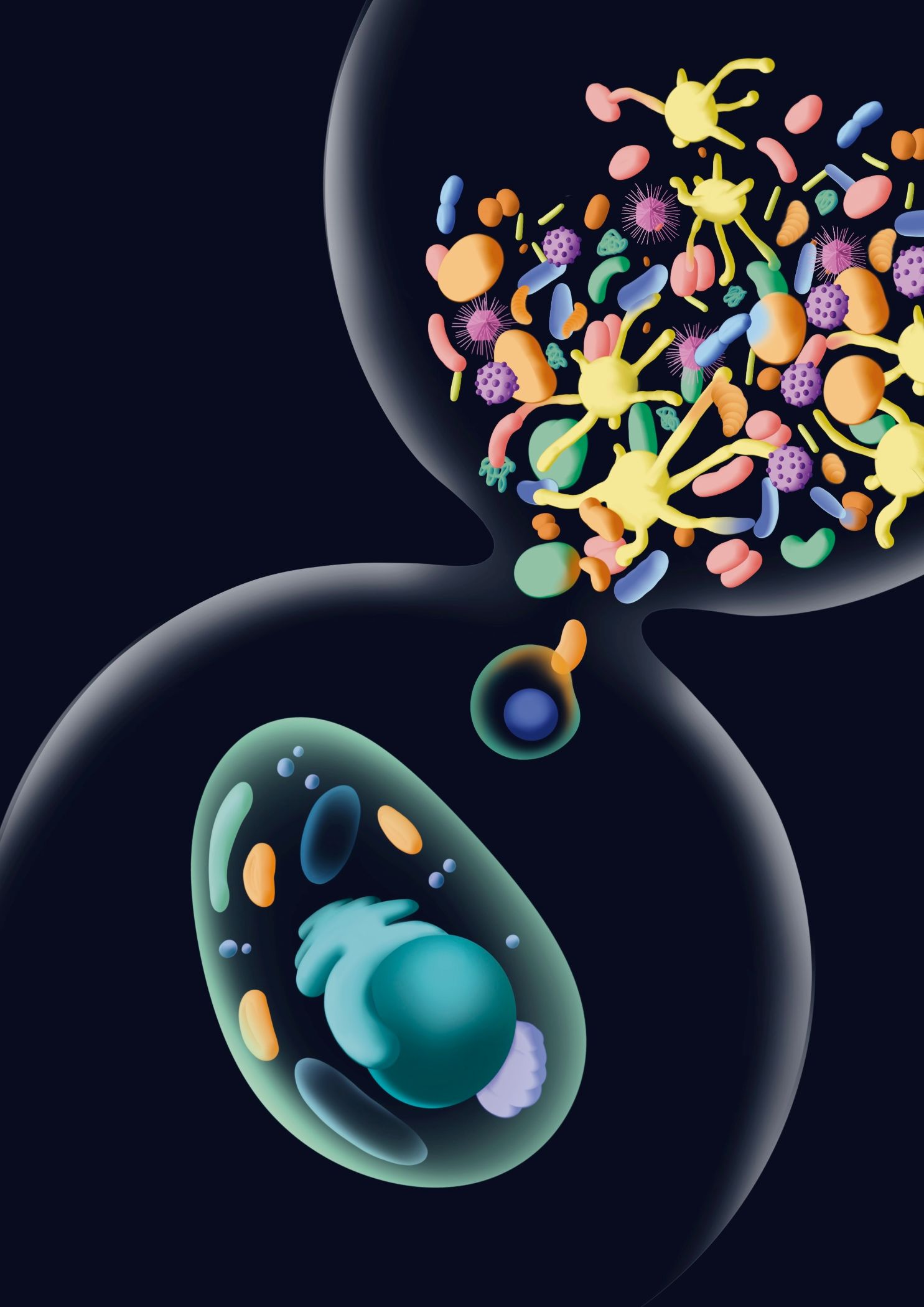
El complejo ecosistema microbiano que facilitó la eucariogénesis se representa mediante la metáfora de un reloj de arena: el pasado es el complejo ecosistema microbiano que da origen a la célula ancestral que engulló a la mitocondria y, posteriormente, al último ancestro común eucariota. / Ilustración: Laura Fraile BSC/IRB.
Alex de Mendoza - eucariotas Nature
Alex de Mendoza
Investigador en epigenómica evolutiva de eucariotas en la Queen Mary University de Londres (Reino Unido)
Este estudio es de gran calidad y toca un tema candente, que es identificar qué fuentes recibió la célula que luego derivó en todas las células con núcleo actuales (los eucariotas), que abarcan animales, hongos o plantas.
La mayor contribución del estudio es identificar varias fuentes bacterianas, más allá de las dos esperables, que son las arqueas de Asgard y la mitocondria. Aquí se proponen otros tres grandes ‘donantes’ de genes, con una contribución similar a la mitocondria a nivel numérico.
Personalmente, la parte que más me gusta es la antes subestimada contribución de los virus gigantes, que son excepcionales por tener muchísimos más genes que otros virus, y con funciones más parecidas a las de vida celular. Esto implica que el proceso de formación de los eucariotas no fue una relación monógama entre dos organismos, sino algo más tipo un ménage à trois con distintos organismos en distintas épocas (¡con enfermedades virales incluidas!), creando un panorama de origen híbrido de nuestro genoma ancestral. Es interesante porque el mensaje de este artículo entra en conflicto con otra publicación también en Nature este mismo año que argumenta lo contrario. Obviamente diferencias metodológicas estarán detrás de las distintas conclusiones, ya que resolver qué pasó hace más de mil millones de años no es tarea fácil, pero yo personalmente creo que este estudio es robusto.
Alfonso Martínez Arias - eucariota Nature
Alfonso Martínez Arias
Profesor de investigación ICREA Senior e investigador en Sistemas de Bioingeniería-MELIS de la Universidad Pompeu Fabra
El trabajo es bueno, original e interesante. El origen de la célula eucariotica es un misterio que solo se ha resuelto parcialmente. La teoría de la unión entre ancestros de la mitocondria y arqueobacterias, de la que fue pionera Lynn Margulis, abrió las puertas para explorar el problema con ideas poco ortodoxas. En lugar de transformaciones graduales clásicas, proponía la existencia de saltos fenotípicos derivados de fusiones entre organismos. Con los años, se han ido acumulando datos en apoyo de esta teoría, pero la complejidad de la célula eucariota indica que esa fusión ancestral no puede dar cuenta de la complejidad.
Este trabajo presenta evidencia sólida y provocativa de varios ciclos de invasiones y adquisiciones parasíticas que, al parecer, predatan la invasión que daría lugar a la mitocondria. Quizás estas invasiones y fusiones prepararon el terreno para otras posteriores. El supuesto papel de los virus en estos procesos evolutivos tempranos es particularmente interesante, pues se sabe que han participado en otros posteriores.
El trabajo es importante no solo porque añade datos importantes sobre el origen de una estructura tan compleja como la célula eucariota, sino porque apoya la noción de procesos no graduales, no darwinianos clásicos, en la emergencia de estructuras y organismos complejos.
Iñaki Ruiz - eucariota Nature
Iñaki Ruiz-Trillo
Investigador principal en el Instituto de Biología Evolutiva (CSIC-UPF)
Mi valoración general del artículo es muy positiva. Se trata de un trabajo técnicamente sólido, basado en un análisis filogenómico masivo, que representa una contribución importante al debate sobre el origen de los eucariotas.
Uno de sus principales méritos es cuestionar la visión excesivamente simplificada de la eucariogénesis como un único evento derivado de una interacción binaria entre una arquea y la futura mitocondria. Además, destaca por la enorme cantidad de genomas analizados. En este sentido, el trabajo encaja bien con propuestas previas que sugerían la participación de múltiples socios microbianos durante la transición hacia los eucariotas, incluyendo posibles contribuciones de grupos como Myxococcales o Planctomycetes.
Sin embargo, creo que algunas de las conclusiones más ambiciosas del artículo deben interpretarse con cautela.
El estudio detecta señales filogenéticas compatibles con contribuciones génicas procedentes de diversos grupos bacterianos, además de las alfa-proteobacterias relacionadas con el origen de la mitocondria. Esto refuerza y amplía observaciones previas. Lo novedoso del trabajo es la escala de los análisis y la integración de estos resultados en un marco más amplio que incorpora también un posible papel para los virus.
Ahora bien, estos resultados no implican necesariamente que podamos identificar con precisión cuáles fueron los socios ecológicos concretos implicados en la eucariogénesis. La razón principal es que la transferencia horizontal de genes es extremadamente frecuente en bacterias. A ello se suman la pérdida génica, la duplicación de genes, la extinción de linajes completos y la dinámica de los pangenomas. Como consecuencia, la historia evolutiva de un gen puede diferir considerablemente de la historia evolutiva de la célula que lo porta.
Por ello, cuando un gen eucariota muestra afinidad con grupos bacterianos actuales como Planctomycetota o Myxococcota, debemos ser prudentes. Lo que probablemente observamos es una relación con un reservorio génico asociado a esos grupos, no necesariamente una evidencia directa de que dichos linajes representen a los donantes originales ni de que existiera una interacción biológica específica con ellos. Es decir, estos genes podrían, pese a su ascendencia a Myxococcales, haber sido parte de la población celular alpha-proteobacteriana que dio lugar a los mitocondrios. La combinación de cromosomas fluidos, pangenomas abiertos y transferencia horizontal masiva puede explicar buena parte de los patrones observados sin necesidad de invocar asociaciones ecológicas concretas. Dicho de otro modo, las mismas señales podrían emerger de redes complejas de intercambio génico dentro de comunidades microbianas donde los genes circulaban entre numerosos organismos durante largos periodos de tiempo.
Aquí surge otra cuestión conceptual: ¿qué entendemos exactamente por LECA?
Tradicionalmente se ha tendido a imaginar LECA [Último Ancestro Común Eucariota] como una célula individual bien definida y creo que los autores lo ven así. Sin embargo, una visión alternativa es considerar LECA como una población diversa, posiblemente con estructura pangenómica, en la que distintos individuos compartían un núcleo común de genes, pero mantenían repertorios accesorios variables.
Si esta visión es correcta, reconstruir la historia de LECA gen por gen puede resultar problemático. Quizá la pregunta más relevante no sea “¿de qué bacteria procede este gen?”, sino “¿qué reservorio génico estaba disponible en las poblaciones ancestrales que dieron lugar a los eucariotas?”. El foco pasaría así de una célula ancestral concreta a una población ancestral dinámica.
Desde esta perspectiva, los resultados del artículo no serían tanto una demostración de asociaciones exclusivas con determinados grupos bacterianos como una confirmación de que la eucariogénesis tuvo lugar en un contexto de intensa conectividad genética dentro de comunidades microbianas complejas.
En resumen, considero que el artículo constituye una aportación importante porque refuerza una visión cada vez más aceptada de la eucariogénesis como un proceso gradual, poblacional y ecológicamente complejo. Sin embargo, debemos ser cautos al interpretar los grupos bacterianos identificados como actores concretos de ese proceso. La transferencia horizontal de genes, la dinámica de los pangenomas y la pérdida génica acumulada durante miles de millones de años limitan nuestra capacidad para reconstruir con precisión quién interactuó con quién en los orígenes de los eucariotas.
Brett J. Baker - eucariota Nature
Brett J. Baker
Profesor asociado en los departamentos de Biología Integrativa y Ciencias Marinas (Instituto de Ciencias Marinas) de la Universidad de Texas en Austin (Estados Unidos)
La posible contribución de virus gigantes a LECA [Último Ancestro Común Eucariota] desde Asgards es, para mí, el aspecto más interesante del estudio. Si bien aún no hemos observado virus gigantes que infecten a Asgards, ni a ninguna otra arquea, creo que hay mucho que aprender al respecto.
En mi opinión, una gran deficiencia del estudio es que los hallazgos parecen estar sesgados por la falta de representación en las bases de datos utilizadas. Por ejemplo, utilizan GTDB de 2022 para el muestreo genómico de procariotas. Esto representa un problema, ya que el número de genomas de Asgards se ha duplicado con creces solo en el último año (véase Appler et al., Nature 2026), que incluye una expansión de Asgards de Heimdall, organismos estrechamente relacionados con los eucariotas.
Edward M. Marcotte - eucariotas Nature
Edward M Marcotte
Profesor de Bioquímica en la Universidad de Texas en Austin (Estados Unidos)
El artículo es técnicamente riguroso y de buena calidad, en general.
El trabajo concuerda con una visión emergente del ancestro unicelular de todos los organismos complejos modernos (denominado LECA, el último ancestro común eucariota) como un microbio nadador con miles de genes y con gran parte de la complejidad de las células modernas ya presente hace entre 1.500 y 1.800 millones de años. Los autores demuestran que los genes de LECA surgieron de múltiples ancestros anteriores, una combinación de genes de diferentes ramas del árbol de la vida microbiana e incluso de virus. Este trabajo, junto con el de otros grupos, nos ofrece una visión de los primeros pasos en la evolución de los organismos complejos modernos.
[En cuanto a posibles limitaciones] Como ocurre con cualquier intento de remontarse tan atrás en el tiempo, incluso si la visión general es correcta, preveo que los detalles seguirán perfeccionándose y cambiando ligeramente con futuros análisis. En particular, a medida que la comunidad científica mundial continúa descubriendo más ramas de la vida microbiana, podemos esperar estimaciones cada vez más precisas del origen de los genes modernos.
Trabajo en la misma área general y, casualmente, mi grupo también publicó un artículo la semana pasada que informa sobre el proteoma y el interactoma de LECA. Nuestro artículo se publicó hace una semana en Cell Genomics. Describe la determinación de los genes codificadores de proteínas (el proteoma) presentes en LECA y luego aborda la cuestión de cómo estas proteínas se organizaron en ‘máquinas moleculares’, capturando la organización física de la maquinaria bioquímica básica en este ancestro crítico de toda la vida compleja moderna. Luego aplicamos esta información para estudiar los organismos actuales, incluyendo el descubrimiento de nuevos genes que afectan, por ejemplo, la densidad ósea o los defectos congénitos, basándonos en estas proteínas e interacciones ancestrales.
En contraste, el artículo del grupo de Toni Gabaldón también define primero los genes en LECA, pero luego aborda la cuestión de su origen, es decir, su ascendencia: ¿surgieron de ancestros bacterianos o arqueales? ¿Se pueden determinar estos orígenes con mayor precisión? Por lo tanto, nuestros dos artículos son bastante complementarios, ya que ambos comparten el mismo objetivo inicial (definir los genes codificadores de proteínas en LECA) y luego los utilizan para plantear preguntas diferentes. No puedo comparar directamente nuestros resultados sin tener acceso a todos los datos (y tiempo para estudiarlos), pero al menos los métodos generales para determinar los genes en LECA parecen bastante comparables (citan nuestra preimpresión de bioRxiv como un método relevante para su artículo), y ambos determinamos un número total similar de familias de genes que nuestros grupos datan de LECA, con el grupo de Gabaldón estimando entre 7.751 y 12.907 familias de genes LECA (ortogrupos) y nuestro artículo estimando entre 6.429 y 10.091 ortogrupos LECA, ambos rangos que dependen del rigor del análisis. Por lo tanto, a grandes rasgos, las primeras partes de nuestros artículos parecen ser muy concordantes.
Así pues, si bien ambos grupos describen los genes de LECA, nuestro trabajo los utiliza como punto de partida para estudiar genes y enfermedades modernas, mientras que el trabajo de Bernabeu, Manzano-Morales, Marcet-Houben y Gabaldón los utiliza para remontarse aún más en el tiempo y estudiar el origen de los genes de LECA.
Bascompte - Eucariota
Jordi Bascompte
Catedrático de Ecología en el departamento de Biología Evolutiva y Estudios Ambientales de la Universidad de Zurich (Suiza)
El origen de la célula compleja es, sin duda, una de las transiciones más importantes en la historia de la vida en nuestro planeta. Sin ella no hubiera sido posible la posterior evolución de los animales pluricelulares como plantas, insectos, anfibios, reptiles y mamíferos, incluyendo a los seres humanos. Es también una de las transiciones más abruptas y enigmáticas. El presente trabajo contribuye a nuestra comprensión de dicha transición proporcionando evidencias muy sólidas de que hubo múltiples eventos de incorporación de componentes genéticos de diversos procariotas incluso antes del famoso evento de endosimbiosis entre la célula huésped, una arquea y el procariota que dio lugar a las mitocondrias.
Estos nuevos resultados abren la puerta a una comprensión más detallada de este importante hito evolutivo. Confirman que el problema continúa siendo relevante a día de hoy y abre las puertas a futuros desarrollos que permitan entender qué tipo de innovaciones en el proceso de regulación genética fueron capaces de lidiar con este origen a partir de múltiples organismos de la célula eucariota. Esencialmente, cómo la vida fue capaz de pasar de un sistema operativo a otro muy diferente, de pasar de una red de regulación genética a una red de redes.
Peretó - Eucariota
Juli Peretó
Catedrático de Bioquímica y Biología Molecular en la Universidad de Valencia
El grupo de Gabaldón publica un estudio robusto, con un gran número de datos y unos métodos filogenómicos avanzados, que permite reconstruir el proteoma del último antepasado común de los eucariotas (LECA), así como analizar las contribuciones de otros linajes a su genoma quimérico.
Más allá de las aportaciones de los dos socios ampliamente reconocidos en el origen de la célula eucariota (una arquea antepasada de las actuales Asgard y una alfa-proteobacteria, la precursora de la mitocondria), debió de haber muchas otras transferencias horizontales de genes desde bacterias y virus en el camino de construcción del genoma de LECA. Los autores identifican al menos dos oleadas de transferencias genéticas hacia el huésped arqueano, antes de la emergencia de la mitocondria.
Además, este estudio aporta hipótesis sobre el contexto ecológico de los tapetes microbianos en el que se debieron producir estos encuentros que resultaron en la complejidad eucariótica.
Las contribuciones de Gabaldón y sus colegas refuerzan la idea de que la eucariogénesis siguió un modo evolutivo genéticamente mestizo (mucho más complejo que el encuentro binario arquea-bacteria) marcado por un tempo dilatado, coherente con sus ideas anteriores sobre un origen tardío de la mitocondria.
Arnau - Eucariota
Arnau Sebé Pedrós
Profesor de investigación ICREA en el Centro de Regulación Genómica (CRG) de Barcelona
Lo que hacen
Los autores tratan uno de los problemas fundamentales de la biología: entender el origen de la célula eucariota. Para ello usan métodos filogenéticos aplicados a datos de genomas actuales, tanto de eucariotas como de procariotas y virus gigantes, con el objetivo de trazar el origen de distintos genes asociados a funciones y estructuras típicamente eucariotas, como el citoesqueleto, la cromatina o el sistema de endomembranas.
Mensaje principal
El resultado más importante es que el estudio refuerza una idea que ya se había sugerido en un artículo anterior de Toni Gabaldón: que la mitocondria (la factoría energética de la célula eucariota) fue una incorporación relativamente tardía. Esto va en contra de una visión más clásica en la que la adquisición temprana de la mitocondria habría sido el motor principal de la evolución de la complejidad eucariota.
Novedad principal
Más allá de datar el origen de la mitocondria a partir de la simbiosis con una alfa-proteobacteria, el estudio también sugiere que la transferencia horizontal de genes desde otras bacterias (y desde virus gigantes) fue un factor clave para explicar el origen de parte de los genes eucariotas. En este sentido, el trabajo propone una visión más compleja y gradual del origen de la célula eucariota, en la que distintas interacciones microbianas habrían contribuido a formar el repertorio génico de los primeros eucariotas. Este sea quizás el mensaje más novedoso de este artículo.
Consistencia con otros resultados recientes
Por último, los resultados están en gran medida de acuerdo, respecto al origen tardío de la mitocondria, con otros dos estudios publicados recientemente en Nature: Kay et al. 2025 y Tobiasson et al. 2026. La principal diferencia es que aquí los autores sugieren una mayor importancia de la transferencia génica horizontal desde bacterias y virus gigantes.
En general me parece un artículo excelente, que ejemplifica a la perfección cómo la secuenciación masiva y el análisis comparado de genomas a lo largo del árbol de la vida están transformando nuestra capacidad para reconstruir, poco a poco, las grandes transiciones evolutivas del pasado.
Snel - Eucariota
Berend Snel
Catedrático en la Universidad de Utrecht (Países Bajos)
La cuestión del origen de la célula eucariota es de una importancia fundamental. Como se ha dicho, “la divergencia básica en la estructura celular, que separa a las bacterias y las cianobacterias del resto de organismos celulares, representa la mayor discontinuidad evolutiva que se puede encontrar en el mundo actual”.
Esta importante cuestión se ha convertido, prácticamente desde el descubrimiento de las asgardarqueas en datos de secuencias metagenómicas, en un campo de investigación muy activo, con numerosos debates y discrepancias. La contribución de Bernabeu et al. analiza una enorme cantidad de genomas diversos mediante un novedoso enfoque computacional para determinar qué genes poseía el ancestro común de los eucariotas y de dónde provenían (es decir, de qué especies). Este artículo demuestra claramente que, a diferencia de modelos anteriores, otras bacterias e incluso quizás virus contribuyeron a los genes del ancestro eucariota. Estudios previos a veces han considerado estas posibles donaciones de genes como ruido filogenético; es decir, la señal real es, por ejemplo, el ancestro mitocondrial bacteriano y la señal observada de ese otro grupo bacteriano es simplemente ruido. Sin embargo, estos autores demuestran de forma muy convincente estas señales de otras bacterias. Demuestran de manera convincente múltiples oleadas de transferencia horizontal de genes.
Además, al determinar el momento en que estos genes fueron donados durante la eucariogénesis, proponen un nuevo escenario sobre cómo las sintrofías posteriores a través de un tapete microbiano podrían haber moldeado el genoma, el metabolismo y la biología celular del protoeucariota. Por ahora se trata simplemente de una hipótesis, pero a diferencia de otras hipótesis en este campo, esta se basa en un análisis a gran escala de datos genómicos. Además, al plantear esta hipótesis, se abre un camino para transformar los datos genómicos en escenarios de sintrofía o refutar hipótesis previamente propuestas.
Conflicto de interés: "He colaborado y publicado con el autor principal del trabajo (Toni Gabaldón)”.
Ratcliff - Eucariota
William C Ratcliff
Profesor e investigador en el departamento de Biología del Instituto Tecnológico de Georgia (Estados Unidos)
Este es un artículo realmente importante. Conozco bien al grupo de Toni Gabaldón (sin conflictos de interés, solo coincidimos en una reunión), y tienen una sólida reputación en este campo. Los análisis parecen meticulosos, y la consistencia de los resultados en múltiples conjuntos de datos es tranquilizadora.
La explicación convencional de la eucariogénesis suele centrarse en un único evento simbiótico: una arquea Asgard incorpora una alfaproteobacteria, esta se convierte en la mitocondria, y a partir de ahí todo se desarrolla. Este trabajo demuestra que la realidad fue mucho más compleja e interesante. El linaje protoeucariota adquirió genes de múltiples grupos bacterianos no relacionados durante un periodo de tiempo probablemente muy extenso y la endosimbiosis mitocondrial, si bien importante, fue solo un episodio en una historia mucho más larga de interacciones. Tendemos a sobrevalorar la mitocondria en nuestra concepción de los orígenes eucariotas, y este artículo corrige esa idea.
Además, tiene una lógica impecable. Si las arqueas de Asgard fueron capaces de establecer una relación simbiótica con un linaje bacteriano, internalizando gran parte de su genoma, no hay razón para suponer que esa fuera la única interacción de este tipo. Estos organismos estaban integrados en complejas comunidades microbianas, y las huellas de esas interacciones ecológicas aún son legibles en los genomas eucariotas miles de millones de años después. Esto también plantea la cuestión de si algunos de estos genes provenían de endosimbiontes anteriores que se perdieron, quizás antes del inicio de la simbiosis que dio origen a la mitocondria. Tendemos a olvidar la ecología cuando analizamos la reconstrucción genómica antigua desde una perspectiva más amplia, y este trabajo nos recuerda su enorme importancia.
El hallazgo del virus es particularmente fascinante. La idea de que los Nucleocytoviricota estuvieran transfiriendo información genética entre linajes dispares y hacia la línea protoeucariota refuerza el creciente reconocimiento de que los virus desempeñan un papel fundamental en la dinámica evolutiva a largo plazo. Los límites entre linajes (¡quizás especialmente para este linaje!) parecen ser mucho más permeables de lo que solemos creer.
En conjunto, el artículo presenta argumentos convincentes de que la eucariogénesis no fue una transición de fase desencadenada por un único evento, sino un proceso gradual de ensamblaje genético a partir de un entorno microbiano diverso, y nos muestra cuánto nos queda por aprender sobre cómo se desarrolló dicho proceso.
Muro - Eucariota
Enrique M. Muro
Investigador principal titular en el Instituto de Evolución Organísmica y Molecular de la Universidad Johannes Gutenberg de Maguncia (Alemania)
La bioinformática continúa iluminando el ‘agujero negro de la biología’ del que surgió la célula eucariota con toda su complejidad. En este trabajo, los autores siguen la estela de Margaret Dayhoff, la madre de la bioinformática que, entre otras contribuciones fundacionales, aportó las primeras evidencias sólidas de un origen alphaproteobacteriano de la mitocondria eucariota. Fue entonces cuando la teoría endosimbiótica de Lynn Margulis ganó la relevancia que tiene hoy en día.
El artículo presentado es técnicamente riguroso y sólido. La hipótesis inicial de que había más procariotas involucradas circulaba entre la comunidad científica, pero este trabajo añade mucha más evidencia e implica a distintos grupos bacterianos, incluso virus gigantes como mediadores en la transferencia génica para que emergiera el primer eucarionte. La consecuencia más relevante que surge de la discusión es que pudo haber una adquisición gradual a través de sucesivos eventos evolutivos. La línea de investigación de mi grupo, que une la biología computacional y la física, apunta a que sucedió algo más, una sinergia donde el todo excedió a la suma de las partes. En ese momento se produjo la transición biológica, creemos que a nivel genético y de forma abrupta.
Desde luego, trabajos como este son una muestra de la capacidad biotecnológica del ser humano. Hace tan solo 25 años nos hubiera parecido inverosímil que se pudieran mostrar evidencias de este tipo (tengamos en cuenta que el origen de la célula eucariota aconteció hace miles de millones de años, nueve ceros en el número de años). La secuenciación y la bioinformática lo están consiguiendo, y algo parecido consigue la paleogenómica, que llega a reconstruir la historia de poblaciones y especies incluso extintas. Sucedió hace tanto tiempo que rebuscar entre esas secuencias parece como buscar entre los posos del café. Pero los autores nos demuestran que todavía nuestra capacidad biotecnológica es válida para encontrar respuestas por esa vía.
- Artículo de investigación
- Revisado por pares
Bernabeu et al.
- Artículo de investigación
- Revisado por pares